An introduction to Susan Pockett: An electromagnetic theory of consciousness
Posted on 1 June 2023 by
Susan Pockett is a neurophysiologist from the University of Auckland, New Zealand. Throughout the past few decades she has published a series of papers on her electromagnetic theory of consciousness – in her own words, that consciousness is identical with certain spatiotemporal patterns in the electromagnetic field. Specifically, it identifies consciousness with the electromagnetic fields surrounding our neurons – the local field potentials – rather than the neurons themselves. What this implies is that what it feels like to be you is what it feels like to be these patterns of electromagnetic fields within the brain.
I originally ran across her work via a blog post by Andrés Gómez Emilsson of the Qualia Research Institute: On Rhythms of the Brain: Jhanas, Local Field Potentials, and Electromagnetic Theories of Consciousness. I wanted to know more, so I found myself reading her 2017 publication, Consciousness Is a Thing, Not a Process. I found her writing to be taut, colourful, and comprehensively researched. I’ll start with a description of the relevant neuroanatomy before summarising her theory.
Consciousness Is a Thing, Not a Process

Abstract: The central dogma of cognitive psychology is ‘consciousness is a process, not a thing’. Hence, the main task of cognitive neuroscientists is generally seen as working out what kinds of neural processing are conscious and what kinds are not. I argue here that the central dogma is simply wrong.
The reader may be aware that the brain is composed of both grey matter and white matter, these being primarily responsible for cognition and connectivity, respectively. If you study a cross-section of the brain, you will see that the grey matter is mostly concentrated in a small layer around 2–4 mm thick on the surface of the cerebral cortex – the neocortex – which in mammals is folded up into gyri and sulci in order to maximise its surface area within the confines of the skull. The cerebral cortex is further subdivided into numerous cortices with various specialised functions. Unfolded, one could imagine the cerebral cortex as like an extremely fine-grained plugboard, with the white matter providing the connections between regions.
Within the neocortex, our pyramidal cells are arranged in parallel with one another into cortical columns, perpendicular to the surface of the cortex:

This surprised me at first, as I’d naïvely imagined a “network” of neurons arranged in a directed graph structure. Instead, we see this radial configuration. For what purpose might it exist?
Ephaptic coupling
When these neurons fire, they produce electric fields – the local field potentials mentioned earlier – which extend beyond the neuronal membrane. As the pyramidal neurons are all aligned with one another like tiny dipoles, if firing in synchrony, then their fields would sum together. As Pockett illustrates:
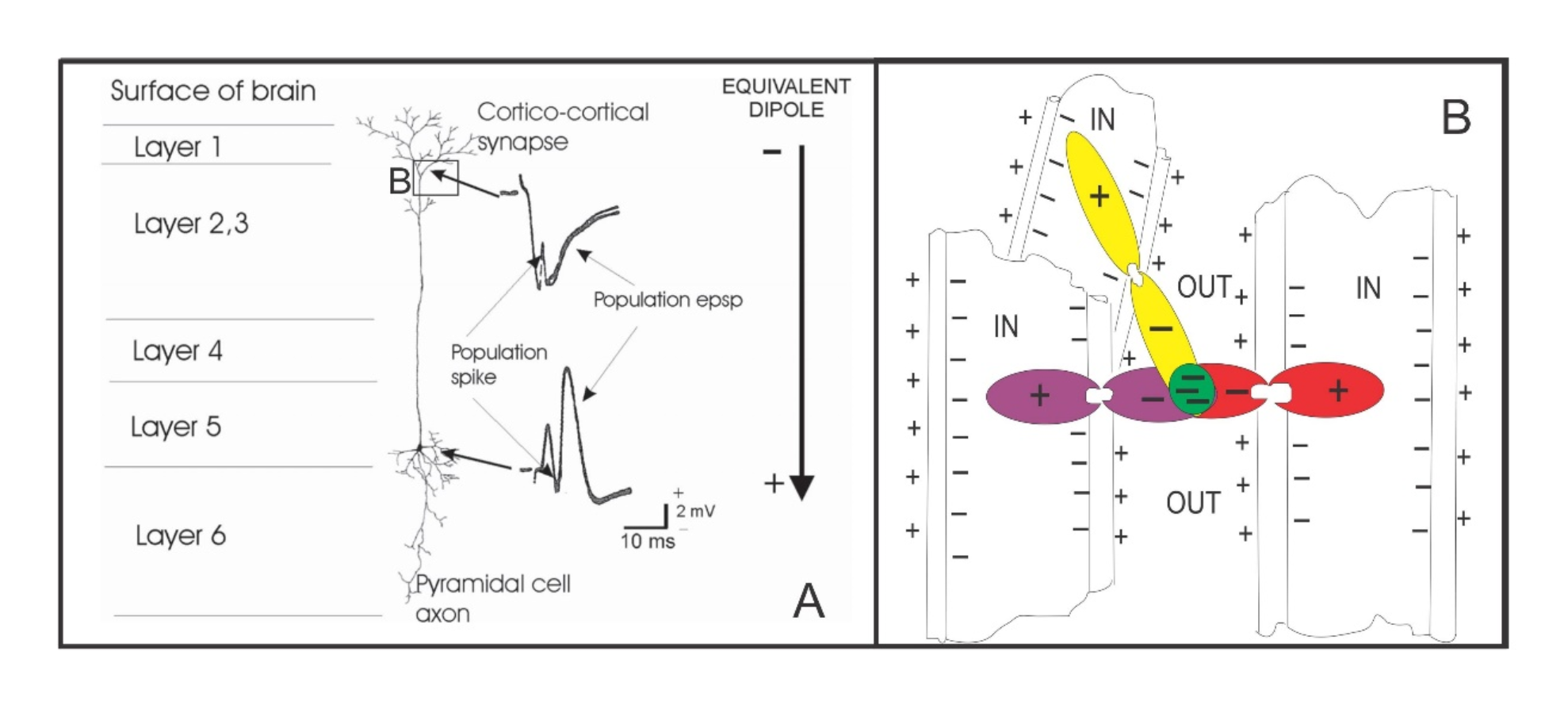
Figure 1. Genesis of local field potentials (LFPs).
(A): LFPs are basically population excitatory postsynaptic potentials (EPSPs) that are generated by synaptic action on the apical dendritic tufts of pyramidal cells. When presynaptic terminals (not shown) release neurotransmitter at site B, positive ions enter the postsynaptic dendrite, leaving a transient negative ‘hole’ in the extracellular fluid. To ‘complete the circuit’ an equal number of positive ions exit the postsynaptic cell nearer the cell body. This process can be modelled as a large equivalent dipole.
(B): Cartoon showing how the negative poles of the smaller dipoles representing entry of positive ions to the postsynaptic dendrites of many synapses can summate to produce the large extracellular voltage transients seen as LFPs (green area). This summation only happens when two conditions are met: (1) many synapses are active synchronously, and (2) the postsynaptic dendrites are anatomically aligned, so that the negative poles of the small dipoles overlap spatially. This explains why large LFPs are not produced during feedforward synaptic activity. Feedforward activity goes mostly through the stellate cells in Layer 4, which do not have anatomically aligned dendrites. Large LFPs are generated only during synchronous feedback synaptic activity, as illustrated in (B).
Next, these fields would evolve over time one way or another, and then influence neuronal firing in return. This is known as ephaptic coupling. Ephaptic coupling represents a closed cycle which would allow consciousness to be non-epiphenomenal – an active influencer of physical events, as opposed to a mere passive observer.
The clinching parallel between consciousness and electromagnetism seems initially to be the observation that consciousness has the ability to act on its own brain. There certainly exists a great deal of evidence that brain-generated EM fields can and do act back on their own brains. This appears to solve the main problem which has always bedeviled philosophical dualism. Traditional substance dualism is routinely rejected on the grounds that it is hard to see how a non-material substance could act on the brain. But here we have a particular kind of non-material substance (electromagnetism) that clearly does act on the brain. Every working neurophysiologist has personal, hands-on experience of making brain tissue respond to experimentally applied EM fields.
To expand upon this concept, Andrés Gómez Emilsson in his Rhythms of the Brain post explains how ephaptic coupling is likely to exert more influence on the phases of neuronal firing than the actual firing thresholds themselves:
As far as I understand the neuroscience literature on LFPs (and ephaptic coupling more generally), the fact that LFPs change firing thresholds is uncontroversial. The question is by “how much”. Most studies find small effects (otoh between 1% and 20% of the variance, but I can look up more precise and recent figures – e.g. see: Ephaptic coupling of cortical neurons).
The more interesting and perhaps significant effect that LFPs have is to change the degree of coherence between neurons. In other words, they may not change much their probability of firing, but do change a lot their probability of firing in phase. You can see how this would lead to interesting self-reinforcing effects.
Andrés also has an evolutionary theory as to what this is all for. By virtue of its electromagnetic field dynamics, the utility of consciousness might lie in its computational properties, which have been recruited by evolution into performing some kind of useful computation that is beneficial to the organism. Perhaps evolution has built a computer out of Maxwell’s equations, and what it feels like to be you, is what it feels like to be this process of computation?
To continue, Pockett details additional aspects of neuroanatomy which would influence the size of these local field potentials and the scope of their influence. She advances the curious notion that not all cortices might be conscious, emphasising the need to find a distinguishing feature between conscious and unconscious areas.
Both psychological experiments on the initiation and control of voluntary actions and direct stimulation of the motor cortex of awake neurosurgical patients combine to support the conclusion that most of the processing underpinning the initiation of actions is unconscious – and more radically, that consciousness usually does not accompany either the initiation or the control of voluntary actions at all. From the point of view of the proposal that conscious experiences are patterned electromagnetic fields, this again suggests that it should be instructive to ask what anatomical feature distinguishes motor areas of cortex. As we have just seen, the motor areas of cortex do not produce conscious experiences, while sensory areas of cortex do. Hence, any major anatomical difference between motor and sensory areas might provide an important clue as to the structural differences between conscious and unconscious EM fields.
Experimental testing
In a self-described manifesto for future experimental testing of the EM field theory of consciousness, Pockett proposes an experimental protocol with which her theory could be verified, using implanted electrodes to neutralise local field potentials without affecting neural activity:
Every scientific theory worth its salt should be falsifiable. One critical experimental test of the idea that conscious experiences are EM patterns was suggested five years ago, but repeated attempts by the author to find a venue in which to carry out the experiment have so far proved unsuccessful. This test involves clamping or zeroing out putatively conscious EM fields using an extracellular version of the classic voltage clamp technique.
The basic aim would be to inject into the extracellular fluid, at a spot predicted by the theory to underpin the simplest possible experimentally inducible experience (for example the sound of a click), sufficient current to just prevent the change of voltage that would normally constitute the local field potential evoked by the auditory stimulus. If this clamp is implemented successfully, it should be possible to compare the current injected by the clamp with the LFP produced by the stimulus in the absence of the clamp, to prove that the clamp does not affect ongoing synaptic activity. If it does not, the current waveform will be a perfect inverse copy of what would normally be the LFP.
To complete the experiment, the patient is then simply asked whether or not they heard a click. If they always hear a click with this particular auditory stimulus in the absence of a clamp, but reliably fail to hear one in the presence of the clamp, the only thing that will have been altered by the clamp is the EM field. The current measurement mentioned above will have ensured than synaptic activity per se – the release of neurotransmitter from the presynaptic terminal, interaction of transmitter with postsynaptic receptors, consequent inflow of ions from the extracellular fluid into the postsynaptic dendrite (potentially leaving a transient ‘hole’ in the extracellular fluid, which hole has here been filled by the injected current) – all continued as normal. So the EM pattern and only the EM pattern has been removed.
We live in an age where recent developments in artificial intelligence have spawned much discussion of what it means to be conscious, but from that field I am yet to see a theory of consciousness that is experimentally verifiable. Here’s one that is.
If an ungrounded metaphysics becomes the dominant stance in an upcoming machine age, the resulting confusion may have unpredictable ethical consequences. The need for a robust theory of consciousness becomes more urgent every day.
In lieu of circular discussion of whether machines might have a subjective experience, why not study something which we do know to be conscious: humans?
Commentary
I find Susan Pockett’s theory to be highly complementary to Steven Lehar’s theory of harmonic resonance, as well as the latest research into geometric eigenmodes. So, if what it feels like to be you is what it feels like to be these patterns of electromagnetic fields within the brain, then one must then imagine a distorted bubble world woven across the folds of the neocortex.
However. I feel that my thoughts are only felt indirectly via nebulous phenomena such as my internal monologue or prophantasia, and that their true representation must reside somewhere outside of the world simulation. My intuition from phenomenological introspection is that it might only be our sensory cortices – visual, auditory, somatosensory, olfactory, and gustatory – that have a direct influence on the state of my world simulation.
How might we weave a world simulation into existence across spatially disparate sensory cortices? It seems like reconciling this with the binding problem would be a big stretch. Perhaps ephaptic coupling in the neocortex is mostly responsible for long-range neural synchrony, and we would benefit by looking for the phenomenally bound worldsim elsewhere – somewhere like the thalamus, as Andrés has suggested?